Dinozaur
Dinozaurii sunt un grup divers de reptile ale cladei Dinosauria. Au apărut pentru prima dată în perioada Triasică, cu 243-233,23 milioane de ani în urmă,[1][2] deși originea exactă și calendarul evoluției dinozaurilor este subiectul cercetării active.[3] Ei au devenit vertebratele terestre dominante după Extincția din Triasic-Jurasic de acum 201 milioane de ani; dominația lor a continuat în Jurasic și Cretacic. Ingineria genetică inversă[4] și înregistrarea fosilă demonstrează că păsările sunt dinozauri moderni cu pene[5] care au evoluat din teropodele primare în timpul Jurasicului Târziu.[6] Ca atare, păsările reprezintă singura linie de dinozauri care a supraviețuit Extincției Cretacic-Paleogen de acum 66 milioane de ani.[7] Astfel, dinozaurii pot fi împărțiți în dinozauri aviari sau păsări; și dinozauri non-aviari.
| Dinozaur | |
|---|---|

| |
| Schelete de diferiți dinozauri non-aviari Sus: Tyrannosaurus, Diplodocus | |
| Clasificare științifică | |
| Regn: | Animalia |
| Încrengătură: | Chordata |
| Subîncrengătură: | Vertebrata |
| Clasă: | Sauropsida |
| Supraordin: | Dinosauria |
| Ordine și Subordine | |
| |
| Modifică text |
|
Dinozaurii sunt un grup variat de animale din punct de vedere taxonomic, morfologic și ecologic. Păsările, cu peste 10.000 de specii vii,[8] sunt cele mai diverse grupuri de vertebrate în afară de pești perciformi.[9] Folosind dovezi fosile, paleontologii au identificat peste 500 de genuri distincte[10] și peste 1.000 de specii diferite de dinozauri non-aviari.[11] Dinozaurii sunt reprezentați pe toate continente de ambele specii existente (păsări) și de resturile fosile.[12] În prima jumătate a secolului XX, înainte ca păsările să fie recunoscute ca fiind dinozauri, majoritatea comunității științifice credea că dinozaurii au fost lenți și cu sânge rece. Majoritatea cercetărilor efectuate încă din anii 1970 au indicat însă că toți dinozaurile erau animale active cu metabolizări ridicate și numeroase adaptări pentru interacțiunea socială. Unii erau erbivori, alții carnivori. Dovezile sugerează că ouarea și construirea cuiburilor sunt trăsături adiționale împărtășite de toți dinozaurii, aviari și non-aviari.
În timp ce dinozaurii erau bipezi ancestrali, multe grupuri dispărute au inclus specii cvadrupede, iar unele erau capabile să treacă de la o poziție la alta. Structurile osoase, cum ar fi coarnele sau crestăturile, sunt comune tuturor grupurilor de dinozauri, iar unele grupuri dispărute au dezvoltat modificări scheletice cum ar fi armura osoasă și spini. În timp ce linia de dinozauri moderni, cei care au reușit să supraviețuiască (păsările), sunt în general, mici din cauza constrângerilor de zbor, mulți dinozauri preistorici (non-aviari și aviari) au avut un corp mare - cei mai mari dinozauri sauropod sunt estimați că au atins lungimi de 39,7 metri [13] și înălțimi de 18 metri[14] fiind cele mai mari animale de uscat ale tuturor timpurilor. Totuși, ideea că dinozaurii non-aviari erau toți gigantici este o concepție greșită, plecată de la ideea că oasele mari sunt mai susceptibile să dureze până când sunt fosilizate. Mulți dinozauri erau destul de mici: Xixianykus, de exemplu, avea o lungime de aproximativ 50 cm.
De la primele fosile de dinozauri descoperite la începutul secolului al XIX-lea, scheletele de dinozauri fosile au fost montate și au reprezentat atracții majore în muzeele din întreaga lume, iar dinozaurii au devenit o parte durabilă a culturii mondiale. Dimensiunile mari ale unor grupuri de dinozauri, precum și natura lor aparent monstruoasă și fantastică, au asigurat apariția regulată a dinozaurilor în cărțile și filmele cel mai bine vândute, precum Jurassic Park. Entuziasmul public persistent pentru animale a dus la o finanțare semnificativă pentru știința dinozaurilor, iar noile descoperiri sunt acoperite în mod regulat de mass-media.
Etimologie
modificareTaxonul "Dinosauria" a fost introdus și definit de paleontologul britanic Richard Owen în 1842, pentru a se referi la "tribul sau sub-ordinul distinct al reptilelor Sauriene".[15] Termenul derivă din două rădăcini grecești δεινός, deinos, însemnând potrivit lui Owen „înspăimântător de mare“, și σαῦρος, sauros însemnând „reptilă“ sau „șopârlă“.[15][16] Owen a ales această combinație lingvistică cu referire la proporțiile impresionante ale acestor animale, precum și la arsenalul lor formidabil de dinți și ghiare.[17]
Alte animale preistorice, inclusiv pterozauri, mosazauri, ihtiozauri, plesiozauri și Dimetrodon, deseori concepute în mod obișnuit ca dinozauri, nu sunt clasificate taxonomic ca dinozauri.[18] Pterozaurii sunt înrudiți de departe cu dinozaurii, fiind membri ai cladei Ornithodira. Celelalte grupuri menționate sunt, ca și dinozaurii și pterozaurii, membri ai Sauropsida (clada reptilelor și păsărilor), cu excepția Dimetrodon (care este un synapsid).
Definiție
modificare
În cadrul nomenclaturii filogenetice, dinozaurii sunt de obicei definiți ca fiind grupul format din cel mai recent strămoș comun (MRCA) al lui Triceratops și al păsărilor și toți descendenții săi.[19] De asemenea, s-a sugerat ca Dinosauria să fie definită cu privire la MRCA de la Megalosaurus și Iguanodon, deoarece acestea erau două din cele trei genuri citate de Richard Owen când a numit Dinosauria.[20] Ambele definiții au ca rezultat aceleași seturi de animale care sunt definite ca dinozauri: "Dinosauria = Ornithischia + Saurischia", cuprinzând anchilozauri (cvadrupede erbivore cu armură), stegosauria (cvadrupede erbivore cu plăci osoase), ceratopsieni (cvadrupede erbivore cu coarne), ornitopozi (erbivore bipede sau cvadrupede inclusând "cioc de rață"), teropode (carnivore bipede și păsări) și sauropodomorfe (erbivore cvadrupede mari cu gât și coadă lungi).[21]
Păsările sunt acum recunoscute ca fiind singura linie supraviețuitoare a dinozaurilor teropode. În taxonomia tradițională, păsările erau considerate o clasă separată care evoluase din dinozauri, un supraordin distinct. Cu toate acestea, majoritatea paleontologilor contemporani interesați de dinozauri resping stilul tradițional de clasificare în favoarea taxonomiei filogenetice; prin această abordare, pentru ca un grup să fie natural, toți descendenții membrilor grupului trebuie să fie incluși și în grup. Astfel, păsările sunt considerate a fi dinozauri, iar dinozaurii nu sunt, prin urmare, dispăruți.[22] Păsările sunt clasificate ca aparținând subgrupului Maniraptora, care sunt coelurosauri, care sunt terapode, care sunt sauriscieni, care sunt dinozauri.[23]
În 2017, cercetările lui Matthew Baron, David B. Norman și Paul M. Barrett au sugerat o revizuire radicală a sistematicii dinozauriene. Analiza filogenetică de către Baron și colab. a considerat Ornithischia ca fiind mai aproape de Theropoda decât Sauropodomorpha, spre deosebire de uniunea tradițională a teropodelor cu sauropodomorfe. Ei au definit clada Ornithoscelida ca referindu-se la grupul care conține Ornithischia și Theropoda. Dinozauria însăși a fost redefinită drept ultimul strămoș comun al Triceratops horridus, Passer domesticus, Diplodocus carnegii și al tuturor descendenților săi, pentru a se asigura că sauropozii și rudele rămân incluse ca dinozauri.[24][25]
Istorie evolutivă
modificareOrigini și evoluția timpurie
modificare
Începând cu mijlocul Trasicului, la aproximativ 20 de milioane de ani după Extincția Permian–Triasic care a distrus aproximativ 95% din toată viața de pe Pământ, dinozaurii s-au desprins din strămoșul lor arhosaur.[26][27] Datarea radiometrică a formațiunii de rocă care conținea fosile din genul de dinozaur timpuriu Eoraptor la vârsta de 231,4 milioane de ani stabilește prezența în înregistrarea fosilelor în acel moment.[28] Paleontologii cred că Eoraptor seamănă cu strămoșul comun al tuturor dinozaurilor;[29] dacă este adevărat, trăsăturile sale sugerează că primii dinozauri erau mici prădători bipezi.[30] Descoperirea ornitodiranilor primitivi, cum ar fi Marasuchus și Lagerpeton, în stratul argentinian de Triasicul Mijlociu, susține acest punct de vedere; analiza fosilelor recuperate sugerează că aceste animale erau într-adevăr niște prădători mici, bipezi. Este posibil ca dinozaurii să fi apărut încă de acum 243 milioane de ani în urmă, după cum reiese din rămășițele din genul Nyasasaurus din acea perioadă, deși fosilele cunoscute ale acestor animale sunt prea fragmentare pentru a spune dacă sunt dinozauri sau rude apropiate dinozaurilor.[31] Recent, s-a stabilit că Staurikosaurus descoperit în formația Santa Maria datează de acum 233,23 milioane de ani, ceea ce îl face mai vechi în epoca geologică decât Eoraptor.[1]
Când au apărut dinozaurii, ei nu au fost animalele terestre dominante. Habitatele terestre erau ocupate de diferite tipuri de arhizauromorfe și therapside, cum ar fi cynodonți și rhynchosauri. Principalii lor concurenți erau pseudosuchia, cum ar fi aetozauri, ornitozoizi și rauisuchieni, care au avut mai mult succes decât dinozaurii.[32] Cele mai multe dintre aceste alte animale au dispărut în Triasic într-una sau două extincții în masă. În prima, cu aproximativ 215 milioane de ani în urmă, o varietate de arhizauromorfe, inclusiv protorosaurii, au dispărut. A urmat Extincția Triasic-Jurasic (cu aproximativ 200 milioane de ani în urmă), când au dispărut majoritatea celorlalte grupuri de arhosauri timpurii, cum ar fi aetozauri, ornitosuchide, fitozauri și rauisuceni. Rhynchosaurii și dicynodonții au supraviețuit (cel puțin în unele zone) cel puțin până în Norianul Târziu sau începutul Rhaetian,[33][34] iar data exactă a dispariției lor este incertă. Aceste pierderi au lăsat loc pentru o faună de crocodilomorfi, dinozauri, mamifere, pterozauri și broaște țestoase.[19] Primele câteva linii ale dinozaurilor timpurii s-au diversificat în etapele Carnian și Norian ale Triasicului, posibil prin ocuparea nișelor grupurilor care au dispărut.[21] De asemenea, în mod deosebit, a existat o rată sporită de dispariție în timpul schimbării climatice majore din Carnian.[35]
Evoluție și paleobiogeografie
modificareEvoluția dinozaurilor după Triasic a fost influențată de schimbările de vegetație și de poziția continentelor. În Triasicul Târziu și Jurasicul Timpuriu continentele erau conectate într-un singur supercontinent numit Pangeea, și a existat o faună comună de dinozauri la nivel mondial compusă mai ales din carnivore Coelophysoidea și erbivore timpurii Sauropodomorpha.[36] Plantele gimnosperme (în special coniferele), s-au răspândit în timpul Triasicului Târziu ca potențială sursă de hrană. Sauropodomorfele timpurii nu aveau mecanisme sofisticate de prelucrare a alimentelor în gură și, prin urmare, trebuie să fi utilizat alte mijloace de a descompune alimentele mai departe de-a lungul tractului digestiv.[37] Omogenitatea generală a faunei dinozauriene a continuat în Jurasicul Mijlociu și Târziu. Printre carnivorii dominanți erau ceratosauria, spinosauroidea și carnosauria, în timp ce printre erbivore, stegosauria și mari sauropode. Dinozaurii din China prezintă unele diferențe, cu teropozi sinraptorid sinraptori specializați și sauropozi neobișnuiți, cu gât lung, Mamenchisaurus.[36] Ankylosaurienii și ornitopozii s-au răspândit tot mai mult, însă prosauropozii au dispărut. Coniferele și alte grupuri de plante, cum ar fi ferigile și coada-calului erau platele dominante. Spre deosebire de prosauropozi timpurii și sauropozi, ornithischienii au dezvoltat mecanisme care să permită procesarea alimentelor în gură, incluzând organe asemăntoare obrajilor pentru a menține mâncarea în gură și mișcări ale fălcilor pentru a mânca alimentele.[37] Un alt eveniment evolutiv notabil al Jurasicului a fost apariția adevăratelor păsări, descendente din Coelurosauria.[38]

În timpul Cretacicului Timpuriu are loc despărțirea continuă a supercontinentului Pangeea, făcând ca fauna dinozaurilor de pe diferite continente să fie tot mai diferențiată. Ankylosaurii, iguanodonții și brachiosauridele s-au răspândit în întreaga Europă, America de Nord și Africa de Nord. Teropode precum marile spinosauride și carcharodontosauride au apărut în Africa, iar în America de Sud au câștigat importanță grupurile de sauropode, cum ar fi rebbachisauridae și titanosauria. În Asia, coelurosaurienii maniraptoran, cum ar fi dromaeosauridele, troodontidele și oviraptorosauria, au devenit teropozii obișnuiți, iar ankylosauridele și ceratopsiile timpurii, ca Psittacosaurus au devenit erbivore importante. Între timp, Australia găzduia o faună a ankylozurieni, a hypsiloponților și a iguanodonților.[36] Stegosaurienii par să fi dispărut la un moment dat la sfârșitul Cretacicul Timpuriu sau la începutul Cretacicului Târziu. O schimbare majoră în Cretacicul Timpuriu, amplificată în Cretacicul Târziu, a fost apariția plantelor cu flori. În același timp, mai multe grupuri de erbivore dinozauri au dezvoltat metode mai sofisticate de a procesa pe cale orală alimentele.

În Cretacicul Târziu au existat trei faune de dinozauri. În America de Nord și nordul Asiei, principalele teropode au fost tiranozauridele și diferitele tipuri de teropozi maniraptoran mai mici, predominante fiind erbivorele hadrosauride, ceratopsieni, anchilozauri și pachycephalosauri. În continentele sudice, care alcătuiseră supercontinentul Gondwana, acum despărțit, abelisauridae erau teropozii cei mai răspândiți și titanosaurienii erau erbivorele dominante. În Europa predominau dromaeosauride, rhabdodonți iguanodonți, anchilozauri, nodosauride și titanosaurienii.[36] Plantele cu flori s-au răspândit foarte mult,[37] și primele ierburi au apărut până la sfârșitul Cretacicului.[39]
Extincția Cretacic-Paleogen, care a avut loc acum aproximativ 66 de milioane de ani, la sfârșitul perioadei Cretacice, a dus la dispariția tuturor grupurilor de dinozauri, cu excepția păsărilor neornithine. Au mai supraviețuit și alte grupuri diapsidice, cum ar fi crocodilienii, sebecosuchia, țestoasele, șopârlele, șerpii, spenodonțienii și choristoderanii.[40]
Liniile supraviețuitoare ale păsărilor neornithine, inclusiv strămoșii ratitelor moderne, ale rațelor și puiilor, și o varietate de păsări de apă, s-au diversificat rapid la începutul Paleogenului, acoperind nișele ecologice lăsate goale de dispariția grupurilor de dinozauri. Se spune adesea că mamiferele au concurat cu neornitinele pentru a domina cele mai multe nise terestre, dar multe dintre aceste grupuri au coexistat cu faune bogate de mamifere pentru cea mai mare parte din Cenozoic.[41]
Clasificare
modificareDinozaurii aparțin unui grup cunoscut sub numele de Archosauria, care include și crocodilienii moderni. În grupul archosauria, dinozaurii se diferențiază cel mai vizibil prin mersul lor. Picioarele dinozaurilor se extind direct sub corp, în timp ce picioarele șopârlelor și crocodilienilor se extind în ambele părți.[42]
Dinozaurii ca o cladă se împart în două ramuri principale: Saurischia și Ornithischia. Saurischia include acei taxoni care împart un strămoș comun mai recent cu păsările decât cu Ornithischia, în timp ce Ornithischia include taxonii care au un strămoș comun mai recent cu Triceratops decât cu Saurischia. Anatomic, aceste două grupuri pot fi distinse cel mai vizibil prin structura lor pelviană. Saurischienii timpurii - "șold de șopârlă", de la cuvintele grecești sauros (σαῦρος), care înseamnă "șopârlă" și ischion (іσχιον) care înseamnă "articulația șoldului" - au în structura șoldului un os pubis orientat înainte.[43] Această formă de bază a fost modificată prin rotirea înapoi a pubisului la diferite grade în mai multe grupuri (Herrerasaurus,[44] therizinosaur,[45] dromaeosauride,[46] și păsări[38]). Saurischia include teropode (exclusiv bipede și cu o varietate largă de diete) și sauropodomorfe (erbivore cu gât lung, cvadrupede).[47][48]
În contrast, Ornithischia - "șold de pasăre", de la cuvintele grecești ornitheios (ὀρνίθειος), care înseamnă "de pasăre" și ischion (ἰσχίον) care înseamnă "articulație șold" - au avut un bazin care seamănă superficial cu pelvisul unei păsări: osul pubis este orientat spre spate. Ornithischia include o varietate de specii care au fost în principal erbivore. (NB: termenii "șold de șopârlă" și "șold de pasăre" sunt termeni improprii - păsările au evoluat din dinozauri cu "șolduri de șopârlă").[42]
-
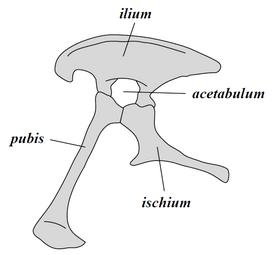 Saurischia - structura pelvisului (partea stângă)
Saurischia - structura pelvisului (partea stângă) -
 Tyrannosaurus - pelvis (arătând structura saurischian – stânga)
Tyrannosaurus - pelvis (arătând structura saurischian – stânga) -
 Ornithischian pelvis structure (partea stângă)
Ornithischian pelvis structure (partea stângă) -
 Edmontosaurus - pelvis (arătând structura ornithischian – stânga)
Edmontosaurus - pelvis (arătând structura ornithischian – stânga)




Taxonomie
modificareMai jos se găsește o clasificare simplificată a grupurilor de dinozauri bazată pe relațiile lor evolutive și organizată pe baza listei speciilor de dinozauri din Mezozoic furnizate de Holtz (2007).[7] O versiune mai detaliată se găsește la clasificarea dinozaurilor. Semnul † este folosit pentru a indica grupuri fără membri vii.
- Dinosauria
- Saurischia („bazin de șopârlă”; include Theropoda și Sauropodomorpha)
- †Herrerasauria (carnivore bipede timpurii)
- Theropoda (toate bipede; majoritatea au fost carnivore)

- †Coelophysoidea (mici, teropode timpurii; include Coelophysis și rude apropiate)
- †Dilophosauridae (teropode timpurii cu creastă și carnivore)
- †Ceratosauria (carnivorele dominante sudice ale Cretacicului)
- Tetanurae ("cozi rigide"; include multe teropode)
- †Megalosauroidea (grup timpuriu de carnivore mari, inclusiv spinosauride semiacvatice)
- †Carnosauria (Allosaurus și rude apropiate, ca Carcharodontosaurus)
- Coelurosauria (teropode cu pene)
- †Compsognathidae (coelurozauri timpurii comuni cu membre anterioare reduse)
- †Tyrannosauridae (Tyrannosaurus și rude apropiate; cu membre anterioare reduse)
- †Ornithomimosauria (similar struțului; fără dinți; carnivore, posibil erbivore)
- †Alvarezsauroidea (mici insectivore cu membre anterioare reduse)
- Maniraptora (membre lungi și degete subțiri)
- †Therizinosauria (erbivore bipede cu ghearele mari la membre și capetele mici)
- †Oviraptorosauria (fără dinți; dieta și stilul de viață sunt incerte)
- †Archaeopterygidae (mici, teropode cu aripi sau păsări primitive)
- †Deinonychosauria (de dimensiune mică sau medie; asemănătoare păsărilor, cu o gheară distinctă)
- Avialae (păsări moderne și rude dispărute)
- †Scansoriopterygidae (înaripate mici primitive cu trei degete lungi)
- †Omnivoropterygidae (înaripate mari, cu coada scurtă)
- †Confuciusornithidae (înaripate fără dinți)
- †Enantiornithes (înaripate zburătoare primitive de copaci)
- Euornithes (păsări zburătoare avansate)
- †Yanornithiformes (păsări cretacice chinezești dințate)
- †Hesperornithes (păsări specializate în scufundări acvatice)
- Aves (păsările moderne, cu cioc și rudele lor dispărute)

- †Sauropodomorpha (erbivore cu capete mici, gâturi lungi, cozi lungi)
- †Guaibasauridae (sauropodomorfe omnivore mici, primitive)
- †Plateosauridae (prosauropode primitive, bipede)
- †Riojasauridae (sauropodomorfe mici, primitive)
- †Massospondylidae (sauropodomorfe mici, primitive)
- †Sauropoda (foarte mari și grele, de obicei peste 15 metri lungime, cvadrupedale)
- †Vulcanodontidae (sauropode primitive)
- †Eusauropoda ("adevăratele sauropode")
- †Cetiosauridae ("reptile balenă")
- †Turiasauria (grupul european de sauropode din Jurasic și Cretacic)
- †Neosauropoda ("noile sauropode")
- †Diplodocoidea (cranii și cozile alungite; dinții în mod obișnuit înguști și în formă de creion)
- †Macronaria (diverse grupuri de sauropode parțial uriașe)
- †Brachiosauridae (gât foarte lung, membrele anterioare sunt mai lungi decât picioarele posterioare)
- †Titanosauria (comun în Cretacicul Târziu în continentele sudice)

- †Ornithischia (diverse erbivore bipede sau cvadrupede)
- †Heterodontosauridae (erbivore/omnivore mai mici cu dinți proeminenți)
- †Thyreophora (dinozauri cu armură osoasă; majoritatea cvadrupede)
- †Ankylosauria (cvadrupede cu armură osoasă)
- †Stegosauria (cvadrupede cu plăci osoase și spini)
- †Neornithischia ("noile ornitopode")
- †Ornithopoda (diverse dimensiuni; bipede și cvadrupede; a dezvoltat o metodă de mestecat folosind flexibilitatea craniului și numeroșii dinți)
- †Marginocephalia (caracterizate de o creștere craniană)
- †Pachycephalosauria ("dinozaur cu capul gros"; bipede )
- †Ceratopsia (cvadrupede cu corn; multe aveau și coarne)
Biologie
modificareCunoștințele despre dinozauri vin din studiul înregistrărilor fosile și nefosile, care includ oase fosilizate, fecale, pene, urme, amprente de piele, țesuturi moi.[49][50] Multe domenii de studiu contribuie la înțelegerea noastră a dinozaurilor, inclusiv fizica (în special biomecanica), chimia, biologia și științele pământului (dintre care paleontologia este o sub-disciplină).[51][52]
Dimensiune
modificare
Dovezile actuale sugerează că mărimea medie a dinozaurilor a variat de-a lungul perioadelor Triasic, Jurasic și Cretacic.[29] Dinozaurii predători teropod, care ocupau cea mai mare parte a carnivorelor terestre din Mezozoic, se încadrau cel mai adesea în categoria 100-1000 kg, atunci când sunt sortați după greutatea estimată în categorii bazate pe ordin de mărime, în timp ce mamiferele predătoare carnivore recente se încadrează în categoria 10-100 kg.[53] Masa corporală a dinozaurului mezozoic este cuprinsă între 1-10 tone.[54] Aceasta contrastează puternic cu mărimea medie a mamiferelor din Cenozoic, estimată de Muzeul Național de Istorie Naturală din Statele Unite, ca fiind între 2-5 kg.[55]
Sauropodele au fost cei mai mari și mai grei dinozauri. Pentru o mare parte a epocii dinozaurilor, cele mai mici sauropode erau mai mari decât orice altceva în habitatul lor, iar cele mai mari aveau un ordin de mărime mai masiv decât orice altceva care a mers de atunci pe Pământ. Mari mamifere preistorice, cum ar fi Paraceratherium (cel mai mare mamifer terestru din toate timpurile), păreau pitici față de gigantele sauropode și doar balenele moderne se apropie sau le depășesc în dimensiune.[56] Există mai multe avantaje pentru mărimea mare a sauropodelor, inclusiv protecția împotriva prădătorilor, reducerea consumului de energie și longevitatea, dar poate că cel mai important avantaj a fost dieta. Animalele mari au o digestie mai eficientă față de animalele mici, deoarece alimentele petrec mai mult timp în sistemul digestiv. Acest lucru le permite să subziste pe baza alimentelor cu o valoare nutritivă mai mică decât la restul animalelor. Rămășițe de sauropode se găsesc în majoritate în formațiuni de roci interpretate ca aparținând unui mediu uscat sau uscat sezonier, iar capacitatea de a mânca cantități mari de cu conținut scăzut de nutrienți ar fi fost avantajoasă în astfel de medii.[14]
Cel mai mare și cel mai mic
modificareOamenii de știință probabil că niciodată nu vor fi siguri de existența celor mai mari și mai mici dinozauri. Acest lucru se datorează faptului că doar un procent mic de animale se fosilizează și cele mai multe dintre acestea rămân îngropate în pământ. Puține exemplare care sunt recuperate sunt schelete complete, iar amprentele de piele și alte țesuturi moi sunt rare. Reconstruirea unui schelet complet prin compararea dimensiunii și morfologiei oaselor cu cele ale unor specii similare, mai bine cunoscute, este o artă inexactă, iar reconstruirea mușchilor și a altor organe este, în cel mai bun caz, un proces de presupunere educată.[57]

Cel mai înalt și mai greu dinozaur cunoscut din schelete este Giraffatitan brancai (anterior clasificat ca o specie de Brachiosaurus). Rămășițele sale au fost descoperite în Tanzania între anii 1907 și 1912. Oase de la mai mulți indivizi cu dimensiuni similare au fost încorporate în scheletul montat și expus la Muzeul de Istorie Naturală din Berlin;[58] acest schelet reconstruit are 12 metri înălțime și 2,5 metri lungime,[59][60] și ar fi aparținut unui animal care a cântărit între 30.000 și 60.000 kilograme. Cel mai lung dinozaur complet este Diplodocus, de 27 metri, descoperit în Wyoming, Statele Unite, și expus în 1907 la Muzeul de Istorie Naturală din Pittsburgh.[61] Cel mai lung dinozaur cunoscut din materiale fosile este Patagotitan: scheletul de la Muzeul American de Istorie Naturală are 37 de metri lungime. Muzeul Carmen Funes din Argentina are un schelet reconstruit de Argentinosaurus cu o lungime de 39,7 metri.[62]

Au existat dinozauri mai mari, dar cunoașterea lor se bazează în întregime pe un număr mic de fosile fragmentare. Cele mai multe dintre cele mai mari exemplare erbivore înregistrate au fost descoperite în anii 1970 sau mai târziu și includ masivul Argentinosaurus, care poate cântări 80.000 până la 100.000 de kilograme și atinge o lungime de 30-40 metri; unii dintre cei mai lungi dinozauri au fost Diplodocus hallorum cu 33,5 metri lungime,[14] Supersaurus cu 33-34 metri[63] și Patagotitan cu 37 metri. Cel mai înalt, Sauroposeidon cu 18 metri înălțime, ar fi putut ajunge la o fereastră la etajul șase. Dinozaurul cel mai greu și cel mai lung e posibil să fi fost Amphicoelias fragillimus, cunoscut numai dintr-un arc nevral parțial, astăzi pierdut, și descris în 1878. Extrapolând din ilustrația acestui os, animalul putea ajuge la o lungime de 58 de metri și să cântărească 122.400 kg.[14] Totuși, nu s-au găsit dovezi suplimentare de sauropode de această dimensiune, iar descoperitorul, Edward Cope, a făcut anterior erori tipografice, ceea ce înseamnă că putem avea o supraestimare în acest caz.[64]
Cel mai mare dinozaur carnivor a fost Spinosaurus, ajungând la o lungime de 12,6-18 metri și cântărind 7-20,9 tone.[65][66] Printre alte teropode mari carnivore se numără: Giganotosaurus, Carcharodontosaurus și Tyrannosaurus.[66] Therizinosaurus și Deinocheirus au fost printre cele mai înalți dintre teropode. Cel mai mare dinozaur Ornithischian a fost, probabil, hadrosauridul Shantungosaurus care a măsurat 16 metri și a cântărit aproximativ 13 tone.
Cel mai mic dinozaur cunoscut este pasărea colibri-albină,[67] cu o lungime de numai 5 cm și o masă de aproximativ 1,8 grame.[68] Cei mai mici dinozauri non-aviari cunoscuți aveau dimensiunea porumbeilor și erau acei teropozi cei mai apropiați de păsări.[69] De exemplu, Anchiornis huxleyi este în prezent cel mai mic dinozaur adult non-aviar descris cu o greutate estimată de 110 grame[70] și o lungime totală a scheletului de 34 cm.[69][70] Printre cei mai mici dinozauri non-aviari erbivori se numără Microceratus și Wannanosaurus, cu o lungime de aproximativ 60 cm fiecare.[7][71]
Comportament
modificare
Multe păsări moderne sunt foarte sociale, adesea trăind în stoluri. Există un consens general asupra faptului că unele comportamente comune la păsări, precum și la crocodili (cele mai apropiate rude vii ale păsărilor), au fost, de asemenea, comune printre grupurile de dinozauri dispărute. Interpretări ale comportamentului dinozaurilor se bazează în general pe poziția scheletului și a habitatului, simularea computerizată a biomecanicii lor și compararea cu animale mderne de nișă ecologică similară.[51] Multe ipoteze despre comportamentul dinozaurilor sunt doar speculative, dar unele găsesc acordul majorității cercetătorilor.
Prima dovadă a comportamentului social al dinozaurilor non-aviari a fost descoperită în 1878 în Bernissart, Belgia, când au fost descoperite 31 de Iguanodon bernissartensis într-o groapă unde se presupune că s-au înecat;[72] totuși, conform ultimelor analize, aceste schelete au fost depozitate în urma a trei evenimente distincte.[73] Au fost descoperite și alte locuri de decese masive. Acestea, împreună cu mai multe urme, sugerează că pentru mai multe specii de dinozauri timpurii comportamentul gregar era obișnuit. Traseele a sute sau chiar de mii de erbivore indică faptul că Hadrosauridele se puteau deplasa în turme mari, la fel ca bizonul american sau antilopa africană. Urme de Sauropod descoperite în Oxfordshire, Anglia, arată faptul că aceste animale au călătorit în grupuri compuse din mai multe specii diferite.[74] Probabil că dinozaurii au format turme pentru a se apăra împotriva prădătorilor, pentru a proteja puii sau pentru a migra periodic. Există dovezi că multe tipuri de dinozauri cu creștere lentă, incluzând diverse teropode, sauropode, anchilozauri, ornitopode și ceratopsieni, au format grupuri ale indivizilor imaturi. Un exemplu este un sit din Mongolia unde s-au găsit rămășițele a peste 20 de Sinornithomimus, cu vârste cuprinse între un an și șapte ani. Această adunare a fost interpretată ca un grup social care a fost prins în noroi.[75] Unii dinozauri carnivori vânau împreună pentru o pradă mai mare.[76][77] Acest stil de viață este mai puțin frecvent la păsările moderne, crocodili și alte reptile, iar dovezile care sugerează vânarea în grup la teropode precum Deinonychus și Allosaurus pot fi interpretate ca fiind rezultatul disputelor fatale între animale care se hrănesc, la fel ca la mulți prădători moderni.[78]

Crestele și alte ornamente osoase ale unor dinozauri ar fi putut fi prea fragile pentru a fi folosite în apărarea activă, considerându-se că erau probabil folosite pentru afișări sexuale sau agresive, deși nu se știe prea multe despre împerecherea dinozaurilor și despre teritorialism. Plăgile de la cap prin mușcături sugerează că, cel puțin, teropodele s-au angajat în confruntări active agresive.[79]
Din punct de vedere comportamental, una dintre cele mai valoroase fosile de dinozauri a fost descoperită în deșertul Gobi în 1971. A inclus un Velociraptor care ataca un Protoceratops,[80] furnizând dovezi că dinozaurii s-au atacat într-adevăr unul pe altul.[81] Dovezi suplimentare sunt o coadă parțial vindecată a unui Edmontosaurus, un dinozaur hadrosaurid; coada este deteriorată în așa fel încât să demonstreze că animalul a fost mușcat de un tiranozar, dar a supraviețuit.[81] Canibalismul printre unele specii de dinozauri a fost confirmat de mărcile dentare găsite în Madagascar în 2003, care implică teropodul Majungasaurus.[82]
Un studiu din 2017 bazat pe liniile de creștere incrementală observate în dinții fosili ai unor dinozauri Ornithischia (Protoceratops și Hypacrosaurus) a concluzionat că incubația ouălor era lungă (3 până la 6 luni), cel puțin la unele specii.[83].
Comparații între inelul scleral de la dinozauri, păsări moderne și reptile au fost folosite pentru a deduce modelele zilnice de activitate ale dinozaurilor. Deși s-a sugerat că cei mai mulți dinozauri erau activi în timpul zilei, aceste comparații au arătat că dinozauri mici de pradă, cum ar fi dromaeosauridele, Juravenator și Megapnosaurus erau ce mai probabil nocturni. Este posibil ca erbivorele mari și medii și omnivorele ca ceratopsia, sauropodomorfe, hadrosauri, ornithomimozauri să fi fost active pe intervale scurte pe parcursul zilei, deși despre Agilisaurus există indicii că a fost diurn.[84]
Comunicare
modificare
Se știe că păsările moderne folosesc pentru comunicare semnale vizuale și auditive, iar diversitatea structurilor ornamentale în rândul dinozaurilor, cum ar fi coarne, creste, gulere și pene, sugerează importanța în biologia dinozaurilor a comunicării vizuale.[85] Reconstrucția culorii penajului la Anchiornis huxleyi, sugerează importanța culorii în comunicarea vizuală la dinozaurii non-aviari.[86] Pe de altă parte, despre abilitățile vocale ale dinozaurilor se știe foarte puțin. Paleontologul Phil Senter crede că dinozaurii non-aviari se bazau în cea mai mare parte pe afișaje vizuale și, eventual, sunete acustice non-vocale, cum ar fi șuieratul, scrâșnirea fălcilor, etc. El afirmă că este puțin probabil ca dinozaurii să fi fost capabil să vocalizeze, deoarece cele mai apropiate rude, crocodilienii și păsările, folosesc diferite mijloace de vocalizare, primul prin laringe și ultimul printr-un siringe, sugerând că au evoluat independent și că strămoșul lor comun a fost mut.[85]
Cele mai timpurii rămășițe ale unui siringe, care are un conținut suficient de minerale pentru fosilizare, au fost găsite într-un specimen Vegavis iaai asemănător cu o rață, datat cu o vechime de 69-66 milioane de ani, iar acest organ este puțin probabil să fi existat la dinozauri non-aviari. Totuși, spre deosebire de Senter, cercetătorii au sugerat că dinozaurii ar fi putut să vocalizeze și că sistemul vocal al păsărilor bazat pe siringe a evoluat dintr-unul bazat pe laringe, mai degrabă decât ca cele două sisteme să fi evoluat independent.[87] Un studiu din 2006 concluzionează că dinozaurii produceau vocalizări cu gura închisă, cum ar fi gânguritul, care apare atât la crocodilieni, cât și la păsări, precum și la alte reptile.
Concluzii recente sugerează că creasta goală a lambeosaurinelor poate funcționa ca o placă de sondare utilizată pentru o varietate largă de vocalizări.[88][89]
Reproducere
modificare
Toți dinozaurii au ouă amniotice cu o carapace dură, produse în mare parte din carbonat de calciu.[90] Majoritatea speciilor creează cuiburi elaborate în formă de castron, cupe, movile, vizuini.[91]Unele specii de păsări moderne nu construiesc cuiburi; o specie de păsări care trăiește pe stânci, Guillemot comun, își depune ouăle pe stânca goală, iar masculul Pinguin imperial are grijă de ou plasându-l între abdomen și picioare. Păsările primitive și mulți dinozauri non-aviari depun adesea ouă în cuiburi comune, masculii incubând în primul rând ouăle. În timp ce păsările moderne au doar un oviduct funcțional și au un singur ou la un moment dat, păsările mai primitive și dinozaurii aveau două oviducte, ca la crocodilii. Unii dinozauri non-aviari, cum ar fi Troodon puteau depozita o pereche de ouă în fiecare zi, sau la două zile, și apoi aveau grijă să eclozeze simultan, amânând incubare până când toate ouăle erau depozitate.[92]

La depunerea ouălor, femelele dezvoltă un tip special de os între osul exterior dur și măduvă. Acest os este bogat în calciu și este folosit pentru a forma coaja oului. Descoperirea acestei proprietăți la un specimen de Tyrannosaurus rex a furnizat dovezi ale osului medular și, pentru prima dată, a permis paleontologilor să stabilească sexul unui specimen de dinozaur fosil. Cercetări ulterioare au descoperit oase medulare la specimene de Allosaurus și Tenontosaurus. Deoarece linia dinozaurilor care include Allosaurus și Tyrannosaurus s-a despărțit de linia care a condus la Tenontosaurus, foarte devreme în evoluția dinozaurilor, acest lucru sugerează că producția de țesut medular este o caracteristică generală a tuturor dinozaurilor.[93]
O altă trăsătură răspândită printre păsările moderne este îngrijirea puilor. Descoperirea unor cuiburi de Maiasaura ("șopârla mamă bună") în Montana în 1978 a arătat că, printre ornitopode, îngrijirea puilor a continuat mult timp după eclozarea ouălor.[94] Este posibil ca acest comportament să fie împărtășit de toți dinozaurii.[94] Un embrion de dinozaur (aparținând probabil Massospondylus) a fost descoperit fără dinți, sugerând astfel că este nevoie de îngrijirea părinților pentru a-l alimenta.[95] Urme găsite pe insula Skye din nord-vestul Scoției au confirmat faptul că ornitopodele își îngrijeau puii.[96]
Toate acestea ar putea explica faptul că unii dinozauri au dezvoltat atât instinct matern, cât și instinct patern, la fel ca la mamiferele și păsările contemporane.
Originea păsărilor
modificarePosibilitatea ca dinozaurii să fie strămoșii păsărilor a fost sugerată pentru prima dată în 1868 de Thomas Henry Huxley.[97] După lucrarea lui Gerhard Heilmann la începutul secolului al XX-lea, teoria păsărilor ca descendenți ai dinozaurilor a fost abandonată în favoarea ideii că păsările sunt descendenți ai Thecodontia; principalul argument care a susținut această teză a fost presupusa lipsă a claviculei la dinozauri.[98] Cu toate acestea, după cum au arătat descoperirile ulterioare, claviculele (sau un singur os creat prin fuzionarea oaselor) nu erau de fapt absente;[38] au fost găsite încă din anul 1924 la Oviraptor, dar au fost identificate greșit drept interclavicul.[99] În anii '70, John Ostrom a reînviat teoria dinozaurilor-păsări,[100] care a câștigat un impuls în deceniile următoare, odată cu apariția analizei cladiste,[101] și o mare creștere în descoperirea micilor teropode și a păsărilor timpurii.[102] De o importanță deosebită au fost fosilele din Formația Yixian, unde au fost găsite o varietate de teropode și păsări timpurii, adesea cu pene de un anumit tip.[5][38] Păsările au în comun peste o sută de trăsături anatomice distincte cu dinozaurii teropod, care acum sunt în general acceptați ca fiind cele mai apropiate rude dispărute.[103] Ele sunt cei mai strâns aliate cu Maniraptora.[38] O minoritate de oameni de știință, dintre care cei mai notabi sunt Alan Feduccia și Larry Martin, au propus alte căi evolutive, inclusiv versiuni revizuite ale propunerii lui Heilmann despre archozaurii primitivi,[104] [77] sau că teropodele maniraptorane sunt strămoșii păsărilor, însă ei înșiși nu sunt dinozauri ci doar convergenți cu dinozaurii.[105]
Pene
modificare
Penele sunt una dintre caracteristicile cele mai recunoscute ale păsărilor moderne și o trăsătură comună cu toate celelalte grupuri de dinozauri. Bazându-se pe distribuția actuală a dovezilor fosile, se pare că penele erau o trăsătură a lor ancestrală, deși una care s-ar fi putut pierde selectiv la unele specii.[106] Dovezi fosile directe de pene sau de structuri asemănătoare cu penele au fost descoperite la o gamă diversă de specii în multe grupuri de dinozauri non-aviari,[5] atât în rândul sauriscienilor cât și al ornitiscienilor. Structuri simple, ramificate, asemănătoare cu penele, sunt cunoscute la heterodontosauride, neornithischia primitive[107] teropode primitive,[108] și ceratopsia primitive. Dovezi pentru pene adevărate, asemănătoare cu cele ale păsărilor moderne, au fost găsite numai în subgrupul teropod Maniraptora, care include: oviraptorozauri, troodontide, dromaeosauride și păsări.[38][109] Structuri tip-pene cunoscute sub denumirea de pycnofibre au fost găsite, de asemenea, la pterosauri.[110]
Archaeopteryx a fost prima fosilă care a dezvăluit o posibilă legătură între dinozauri și păsări. Este considerată o fosilă de tranziție, prin faptul că prezintă caracteristici ale ambelor grupuri. A apărut la lumină la doar doi ani după Originea speciilor a lui Darwin, descoperirea ei stimulând dezbateri între susținătorii biologiei evoluționiste și cei ai creaționismului. Această pasăre timpurie este atât de asemănătoare cu un dinozaur, încât fără o amprentă clară a penei din piatra înconjurătoare, cel puțin un specimen a fost confundat cu Compsognathus.[111] Începând cu anii 1990, au fost descoperiți o serie de dinozauri cu pene, oferind dovezi mai puternice despre relația strânsă dintre dinozauri și păsările moderne. Cele mai multe dintre aceste specimene au fost descoperite în Formația Yixian, Liaoning, din nord-estul Chinei, care făcea parte dintr-un continent insular în timpul Cretacicului. Lipsa unor dovezi fosilizate pe scară largă pentru dinozauri non-aviari cu pene se poate datora structurilor delicate cum ar fi pielea și penele, care rareori sunt conservate prin fosilizare și, prin urmare, sunt absente în registrul fosil.[112]
Descrierea dinozaurilor cu pene nu a fost lipsită de controverse; probabil, cei mai vocali critici au fost Alan Feduccia și Theagarten Lingham-Soliar, care au propus că unele fosile de tip pene sunt rezultatul descompunerii fibrelor colagene subcutanate ale dinozaurilor[113][114] [115] și că dinozaurii maniraptoran cu pene paletate nu erau de fapt dinozauri, ci convergenți cu dinozaurii.[105] [40][114] Totuși, opiniile lor nu au fost în mare parte acceptate de alți cercetători, ajungând până la punctul în care natura științifică a propunerilor lui Feduccia a fost pusă la îndoială.[116] [111]
În 2016, s-a raportat că o coadă de dinozaur cu pene a fost găsită într-un chihlimbar. Fosila are o vechime de aproximativ 99 de milioane de ani.[5][117][118]
Schelet
modificareDeoarece penele sunt adesea asociate cu păsările, dinozaurii cu pene sunt adesea descriși ca o verigă lipsă între păsări și dinozauri. Trăsături multiple ale scheletului, comune între cele două grupuri, reprezintă o altă linie importantă de dovezi pentru paleontologi. Zone ale scheletului cu asemănări importante includ gâtul, pubisul, oasele carpului, braț și centura pectorală, furcula (os) și osul de la piept. Comparând scheletul păsărilor și al dinozaurilor prin analiza cladistică se întărește această legătură.[119]
Istoria studiilor
modificareFosile de animale mari mezozoice sunt cunoscute încă de la începuturile omenirii, dar adevărata lor identificare a apărut după interpretări mitologice de milenii, și după decenii de ipoteze fanteziste. Chinezii i-au considerat a fi oase de dragon și le-a documentat ca atare. De exemplu, Hua Yang Guo Zhi, o carte scrisă de Chang Qu în timpul dinastiei Jin occidentale (265-316), a raportat descoperirea oaselor de dragon la Wucheng în provincia Sichuan.[120] Țăranii din centrul Chinei au descoperit de mult oasele fosilizate ale "dragonilor" și le-au utilizat în medicamentele tradiționale. În Europa, fosilele dinozaurilor erau, în general, considerate a fi rămășițele unor giganți și a altor creaturi biblice.[121]
Descrierile științifice despre ceea ce ar fi recunoscute în prezent ca oase de dinozaur au apărut pentru prima dată la sfârșitul secolului al XVII-lea în Anglia. O parte a unui os, acum cunoscut ca fiind femurul unui Megalosaurus,[122] a fost recuperat dintr-o carieră de calcar de la Cornwell, în apropiere de Chipping Norton, Oxfordshire, în 1676. Fragmentul a fost trimis lui Robert Plot, profesor de chimie la Universitatea din Oxford și primul curator al Muzeului Ashmolean, care a publicat o descriere în "Istoria Naturală a Oxfordshire" în 1677. A identificat corect osul ca extremitatea inferioară a femurului unui animal mare și a recunoscut că era prea mare pentru a aparține oricărei specii cunoscute. De aceea, el a concluzionat că este osul coapsei unui om gigantic similar cu cei menționați în Biblie. În 1699, Edward Lhuyd, un prieten al lui Sir Isaac Newton, a fost responsabil pentru primul tratament științific publicat despre ceea ce în prezent este recunoscut ca dinozaur atunci când a descris și a numit un dinte de sauropod, Rutellum implicatum[123][124] care fusese găsită în Caswell, lângă Witney, Oxfordshire.

Între anii 1815 și 1824, William Buckland, profesor de geologie la Oxford, a adunat mai multe oase fosilizate de Megalosaurus și a devenit prima persoană care a descris un dinozaur într-un jurnal științific.[122][125] Al doilea gen de dinozaur identificat, Iguanodon, a fost descoperit în 1822 de către Mary Ann Mantell - soția geologului englez Gideon Mantell. Gideon Mantell a recunoscut asemănările dintre fosilele sale și oasele iguanelor moderne. El a publicat concluziile sale în 1825.[126][127] [98]
Studiul acestor "fosile de mari șopârle" a căpătat curând un interes deosebit printre oamenii de știință europeni și americani, iar în 1842 paleontologul englez Richard Owen a inventat termenul "dinozaur". El a recunoscut că rămășițele care au fost găsite până acum, Iguanodon, Megalosaurus și Hylaeosaurus, aveau în comun anumite trăsături și a decis să le prezinte ca un grup taxonomic distinct. Cu sprijinul printului Albert, soțul reginei Victoria, Owen a înființat Muzeul de Istorie Naturală din Londra pentru a afișa colecția națională de fosile de dinozaur și alte expoziții biologice și geologice.[128]
Între anii 1815 și 1824, William Buckland, profesor de geologie la Oxford, a adunat mai multe oase fosilizate de Megalosaurus și a devenit prima persoană care a descris un dinozaur într-un jurnal științific.[122][125] Al doilea gen de dinozaur identificat, Iguanodon, a fost descoperit în 1822 de către Mary Ann Mantell - soția geologului englez Gideon Mantell. Gideon Mantell a recunoscut asemănările dintre fosilele sale și oasele iguanelor moderne. El a publicat concluziile sale în 1825.[126][127]
Studiul acestor "fosile de mari șopârle" a căpătat curând un interes deosebit printre oamenii de știință europeni și americani, iar în 1842 paleontologul englez Richard Owen a inventat termenul "dinozaur". El a recunoscut că rămășițele care au fost găsite până acum, Iguanodon, Megalosaurus și Hylaeosaurus, aveau în comun anumite trăsături și a decis să le prezinte ca un grup taxonomic distinct. Cu sprijinul printului Albert, soțul reginei Victoria, Owen a înființat Muzeul de Istorie Naturală din Londra pentru a afișa colecția națională de fosile de dinozaur și alte expoziții biologice și geologice.[128]



În 1858, William Parker Foulke a descoperit primul dinozaur american cunoscut, în gropi de marnă în micul oraș Haddonfield, New Jersey. (Deși fosilele fuseseră descoperite înainte, natura lor nu fusese corect deslușită) Creatura a fost numită Hadrosaurus foulkii. A fost o descoperire extrem de importantă: Hadrosaurus a fost unul dintre primele schelete aproape complete de dinozaur (primul a fost în 1834, în Maidstone, Anglia), și era clar o creatură bipedă. Aceasta a fost o descoperire revoluționară deoarece, până în acel moment, mulți oameni de știință credeau că dinozaurii mergeau pe patru picioare, la fel ca șopârlele. Descoperirile lui Foulke au declanșat un val de dinozaur manie în Statele Unite.[129]
Dinosaur mania a fost exemplificată de rivalitatea feroce dintre Edward Drinker Cope și Othniel Charles Marsh, ambii încercând să fie primii în descoperirea de noi dinozauri, în ceea ce a devenit cunoscut sub numele de Războiul oaselor. Probabil că învrăjbirea a apărut atunci când Marsh a subliniat în mod public că reconstrucția lui Cope a unui schelet de Elasmosaurus fost eronată: Cope a plasat din greșeală capul plesiosaurului unde ar fi trebuit să fie coada. Lupta dintre cei doi oameni de știință a durat peste 30 de ani, terminând în 1897, când Cope a murit după ce și-a risipit toată averea pe vânătoarea dinozaurilor. Marsh 'a câștigat' concursul în primul rând pentru că a fost mai bine finanțat printr-o relație cu Studiul geologic american. Din păcate, multe specimene de dinozaur valoroase au fost deteriorate sau distruse din cauza metodelor celor doi: de exemplu, excavatoarele lor foloseau adesea dinamită pentru a scoate la iveală oasele. În ciuda metodelor lor nerafinate, contribuțiile lui Cope și Marsh la paleontologie au fost uriașe: Marsh a descoperit 86 de specii noi de dinozaur și Cope a descoperit 56, un total de 142 de specii noi. Colecția lui Cope se află în prezent la Muzeul American de Istorie Naturală din New York, în timp ce colecția lui Marsh este expusă la Muzeul de istorie naturală Peabody de la Universitatea Yale.[130][7]
După 1897, căutarea fosilelor dinozaurilor s-a extins pe toate continentele, inclusiv în Antarctica. Primul dinozaur antarctic care a fost descoperit, anchilozaurul Antarctopelta oliveroi, a fost găsit pe insula James Ross în 1986,[131]
"Zona fierbinte" a actualelor fosile de dinozauri includ sudul Americii de Sud (în special Argentina) și China. China, în special, a produs numeroase specimene de dinozaur cu pene excepționale datorită geologiei unice, precum și a climei aride din vechime care a favorizat fosilarea.[112]
Țesuturi moi și ADN
modificare
Unul dintre cele mai bune exemple de urme de țesuturi moi într-un dinozaur fosil a fost descoperit la Pietraroia, Italia. Descoperirea a fost raportată în 1998 și a descris specimenul unui coelurosaur mic și foarte tânăr, Scipionyx samniticus. Fosile includ porțiuni ale intestinelor, colonului, ficatului, mușchilor și traheelor acestui dinozaur imatur.[49]
În ediția din martie 2005 a revistei Science, paleontologul Mary Higby Schweitzer și echipa ei au anunțat descoperirea unui material flexibil care seamănă cu țesuturi moi în interiorul unui os Tyrannosaurus rex de 68 de milioane de ani din formațiunea Hell Creek din Montana. După recuperare, țesutul a fost rehidratat de echipa de știință.[50] Atunci când osul fosilizat a fost tratat timp de câteva săptămâni pentru a elimina conținutul mineral din cavitatea fosilă a măduvei osoase (un proces numit demineralizare), Schweitzer a găsit dovezi ale structurilor intacte, cum ar fi vase sanguine, matricea osoasă și țesut conjunctiv (fibre osoase). Examinarea sub microscop a relevat în continuare că presupusul țesut moale de dinozaur a păstrat structuri fine (microstructuri) chiar la nivel celular. Natura exactă și compoziția acestui material și implicațiile descoperirii lui Schweitzer nu sunt încă clare.[50]
În 2009, o echipă care l-a inclus pe Schweitzer, a anunțat că, folosind o analiză mai atentă, ei și-au dublat rezultatele prin găsirea unui țesut moale similar într-un dinozaur-cioc-de-rață, Brachylophosaurus canadensis, găsit în Formația Râul Judith din Montana. Aceasta conținea țesuturi mai detaliate, până la celule osoase conservate. Printre alte materiale găsite în os au fost colagenul, ca și la osul Tyrannosaurus. Tipul de colagen pe care un animal îl are în oasele sale variază în funcție de ADN-ul său și, în ambele cazuri, acest colagen a fost de același tip găsit la struți și la găinile moderne.[132]
În două cazuri diferite s-a raportat extracția ADN-ului vechi din fosilele dinozaurilor,[133] însă, în urma verificărilor și evaluărilor nici unul dintre aceste rapoarte nu a putut fi confirmat.[134] Totuși, o peptidă funcțională implicată în construirea unei viziuni teoretice a unui dinozaur a fost dedusă prin metode analitice de reconstrucție filogenetică asupra secvențelor ADN de la specii strâns înrudite, cum ar fi reptilele și păsările.[135] În plus, mai multe proteine, inclusiv hemoglobina,[136] au fost prezumptiv detectate în fosile de dinozaur.[137][138]
În 2015, cercetătorii au raportat că au găsit structuri similare celulelor sanguine și fibrelor de colagen, conservate în fosilele osoase a șase exemplare de dinozaur din Cretacic, care au o vechime de aproximativ 75 de milioane de ani.[139][140]
Vezi și
modificareReferințe și note
modificare- ^ a b Langer, M.C.; Ramezani, J.; Da Rosa, Á.A.S. (). „U-Pb age constraints on dinosaur rise from south Brazil”. Gondwana Research. X (18): 133–140. Bibcode:2018GondR..57..133L. doi:10.1016/j.gr.2018.01.005. ISSN 1342-937X.
- ^ Alcobar, Oscar A.; Martinez, Ricardo N. (). „A new herrerasaurid (Dinosauria, Saurischia) from the Upper Triassic Ischigualasto Formation of northwestern Argentina”. ZooKeys. 63 (63): 55–81. doi:10.3897/zookeys.63.550. PMC 3088398
 . PMID 21594020.
. PMID 21594020.
- ^ Nesbitt, Sterling J.; Barrett, Paul M.; Werning, Sarah; Sidor, Christian A.; Charig, Alan J. (). „The oldest dinosaur? A Middle Triassic dinosauriform from Tanzania”. Biology Letters. 9 (1): 20120949. doi:10.1098/rsbl.2012.0949. PMC 3565515 . PMID 23221875.
- ^ „The Dino-Chickens Are Coming”.
- ^ a b c d St. Fleur, Nicholas (). „That Thing With Feathers Trapped in Amber? It Was a Dinosaur Tail”. The New York Times. Accesat în .
- ^ Lee, MichaelS.Y.; Cau, Andrea; Naish, Darren; Dyke, Gareth J. (). „Sustained miniaturization and anatomical innovation in the dinosaurian ancestors of birds”. Science. 345 (6196): 562–566. Bibcode:2014Sci...345..562L. doi:10.1126/science.1252243. PMID 25082702. Accesat în .
- ^ a b c d Holtz, Thomas R. Jr. (). Dinosaurs: the most complete, up-to-date encyclopedia for dinosaur lovers of all ages. New York: Random House. ISBN 978-0-375-82419-7.
- ^ „Numbers of threatened species by major groups of organisms (1996–2012)” (PDF). IUCN Red List. . Arhivat din original (PDF) la . Accesat în .
- ^ Alfaro, M.E.; F. Santini, C. Brock; H. Alamillo, A. Dornburg. D.L. Rabosky; G. Carnevale, L.J. Harmon (). „Nine exceptional radiations plus high turnover explain species diversity in jawed vertebrates”. Proceedings of the National Academy of Sciences of the United States of America. 106 (32): 13410–13414. Bibcode:2009PNAS..10613410A. doi:10.1073/pnas.0811087106. PMC 2715324 . PMID 19633192.
- ^ Wang, S.C.; Dodson, P. (). „Estimating the Diversity of Dinosaurs”. Proceedings of the National Academy of Sciences of the United States of America. 103 (37): 13601–13605. Bibcode:2006PNAS..10313601W. doi:10.1073/pnas.0606028103. PMC 1564218 . PMID 16954187.
- ^ Amos J (). „Will the real dinosaurs stand up?”. BBC News. Accesat în .
- ^ MacLeod, N; Rawson, PF; Forey, PL; Banner, FT; Boudagher-Fadel, MK; Bown, PR; Burnett, JA; Chambers, P; Culver, S; Evans, SE; Jeffery, C; Kaminski, MA; Lord, AR; Milner, AC; Milner, AR; Morris, N; Owen, E; Rosen, BR; Smith, AB; Taylor, PD; Urquhart, E; Young, JR (). „The Cretaceous–Tertiary biotic transition”. Journal of the Geological Society. 154 (2): 265–292. Bibcode:1997JGSoc.154..265M. doi:10.1144/gsjgs.154.2.0265.
- ^ Sellers, William Irvin; Margetts, Lee; Coria, Rodolfo Aníbal; Manning, Phillip Lars (). „March of the Titans: The Locomotor Capabilities of Sauropod Dinosaurs”. PLOS One. 8 (10): e78733. Bibcode:2013PLoSO...878733S. doi:10.1371/journal.pone.0078733. ISSN 1932-6203. PMC 3864407 . PMID 24348896.

- ^ a b c d Carpenter, Kenneth (). „Biggest of the big: a critical re-evaluation of the mega-sauropod Amphicoelias fragillimus”. În Foster, John R.; Lucas, Spencer G. Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin 36. Albuquerque: New Mexico Museum of Natural History and Science. pp. 131–138.
- ^ a b Owen, Richard (). „Report on British fossil reptiles. Part II.”. Report of the Eleventh Meeting of the British Association for the Advancement of Science; Held at Plymouth in July 1841. London: 60–204. From p. 103: "The combination of such characters … will, it is presumed, be deemed sufficient ground for establishing a distinct tribe or sub-order of Saurian Reptiles, for which I would propose the name of Dinosauria*. (*Gr. δεινός, fearfully great; σαύρος, a lizard. … )"
- ^ „Liddell–Scott–Jones Lexicon of Classical Greek”. Accesat în .
- ^ Farlow, J.O.; Brett-Surman, M.K. (). „Preface”. În Farlow, J.O.; Brett-Surman, M.K. The Complete Dinosaur. Indiana University Press. pp. ix–xi. ISBN 978-0-253-33349-0.
- ^ Chamary, JV (). „Dinosaurs, Pterosaurs And Other Saurs – Big Differences”. Forbes. Accesat în .
- ^ a b Benton, Michael J. (). „Origin and relationships of Dinosauria”. În Weishampel, David B.; Dodson, Peter; Osmólska, Halszka. The Dinosauria (ed. 2nd). Berkeley: University of California Press. pp. 7–19. ISBN 978-0-520-24209-8.
- ^ Olshevsky, G. (). „An annotated checklist of dinosaur species by continent”. Mesozoic Meanderings. 3: 1–157.
- ^ a b Langer, Max C.; Martin D. Ezcurra; Jonathas S. Bittencourt; Fernando E. Novas (). „The origin and early evolution of dinosaurs”. Biological Reviews. 85 (1): 65–66, 82. doi:10.1111/j.1469-185x.2009.00094.x. PMID 19895605.
- ^ „Using the tree for classification”. University of Berkeley.
- ^ Padian, K. (2004). "Basal Avialae". In Weishampel, D.B.; Dodson, P.; Osmolska, H. (eds.). The Dinosauria (Second ed.). Berkeley: University of California Press. pp. 210–231. ISBN: 0-520-24209-2
- ^ Wade, Nicholas (). „Shaking Up the Dinosaur Family Tree”. The New York Times. Accesat în .
- ^ Baron, Matthew G.; Norman, David B.; Barrett, Paul M. (). „A new hypothesis of dinosaur relationships and early dinosaur evolution”. Nature. 543 (7646): 501–506. Bibcode:2017Natur.543..501B. doi:10.1038/nature21700. PMID 28332513. Accesat în .
- ^ Kump LR; Pavlov A; Arthur MA (). „Massive release of hydrogen sulfide to the surface ocean and atmosphere during intervals of oceanic anoxia”. Geology. 33 (5): 397–400. Bibcode:2005Geo....33..397K. doi:10.1130/G21295.1.
- ^ Tanner LH; Lucas SG; Chapman MG (). „Assessing the record and causes of Late Triassic extinctions” (PDF). Earth-Science Reviews. 65 (1–2): 103–139. Bibcode:2004ESRv...65..103T. doi:10.1016/S0012-8252(03)00082-5. Arhivat din original (PDF) la . Accesat în .
- ^ Alcober, Oscar A.; Martinez, Ricardo N. (). „A new herrerasaurid (Dinosauria, Saurischia) from the Upper Triassic Ischigualasto Formation of northwestern Argentina”. ZooKeys. 63 (63): 55–81. doi:10.3897/zookeys.63.550. PMC 3088398 . PMID 21594020.
- ^ a b Sereno PC (). „The evolution of dinosaurs”. Science. 284 (5423): 2137–2147. doi:10.1126/science.284.5423.2137. PMID 10381873.
- ^ Sereno, P.C.; Forster, Catherine A.; Rogers, Raymond R.; Monetta, Alfredo M. (). „Primitive dinosaur skeleton from Argentina and the early evolution of Dinosauria”. Nature. 361 (6407): 64–66. Bibcode:1993Natur.361...64S. doi:10.1038/361064a0.
- ^ Nesbitt, S.J., Barrett, P.M., Werning, S., Sidor, C.A., and A.J. Charig. (2012). "The oldest dinosaur? A Middle Triassic dinosauriform from Tanzania." Biology Letters.
- ^ Stephen L. Brusatte (). „Superiority, Competition, and Opportunism in the Evolutionary Radiation of Dinosaurs”. Science. 321 (5895): 1485–1488. doi:10.1126/science.1161833. PMID 18787166.
- ^ Justin A. Spielmann; Spencer G. Lucas; Adrian P. Hunt (). „The first Norian (Revueltian) rhynchosaur: Bull Canyon Formation, New Mexico, U.S.A” (PDF). New Mexico Museum of Natural History and Science Bulletin. 61: 562–566. Arhivat din original (PDF) la . Accesat în .
- ^ Tomasz Sulej; Grzegorz Niedźwiedzki (). „An elephant-sized Late Triassic synapsid with erect limbs”. Science. in press. doi:10.1126/science.aal4853. PMID 30467179.
- ^ „Fossil tracks in the Alps help explain dinosaur evolution”. The Economist (în engleză). Accesat în .
- ^ a b c d Holtz, Thomas R. Jr.; Chapman, Ralph E.; Lamanna, Matthew C. (). „Mesozoic biogeography of Dinosauria”. În Weishampel, David B.; Dodson, Peter; Osmólska, Halszka. The Dinosauria (ed. 2nd). Berkeley: University of California Press. pp. 627–642. ISBN 978-0-520-24209-8.
- ^ a b c Fastovsky, David E.; Smith, Joshua B. (). „Dinosaur paleoecology”. În Weishampel, David B.; Dodson, Peter; Osmólska, Halszka. The Dinosauria (ed. 2nd). Berkeley: University of California Press. pp. 614–626. ISBN 978-0-520-24209-8.
- ^ a b c d e f Padian K (). „Basal avialae”. The Dinosauria (2d edition). University of California Press. pp. 210–231. ISBN 978-0-520-24209-8.
- ^ Prasad, V.; Strömberg, CA; Alimohammadian, H; Sahni, A (). „Dinosaur coprolites and the early evolution of grasses and grazers”. Science. 310 (5751): 1170–1180. Bibcode:2005Sci...310.1177P. doi:10.1126/science.1118806. PMID 16293759.
- ^ a b Archibald, J. David; Fastovsky, David E. (). „Dinosaur Extinction”. În Weishampel, David B.; Dodson, Peter; Osmólska, Halszka. The Dinosauria (ed. 2nd). Berkeley: University of California Press. pp. 672–684. ISBN 978-0-520-24209-8.
- ^ Lindow, B.E.K. (2011). "Bird Evolution Across the K–Pg Boundary and the Basal Neornithine Diversification." In Dyke, G. and Kaiser, G. (eds.)Living Dinosaurs: The Evolutionary History of Modern Birds, John Wiley & Sons, Ltd, Chichester, UK. doi:10.1002/9781119990475.ch14
- ^ a b Brusatte, Stephen L. (). Dinosaur Paleobiology (ed. 1.). New York: Wiley, J. pp. 9–20, 21. ISBN 978-0-470-65658-7.
- ^ Benton, M.J. (). Vertebrate Paleontology. Blackwell Publishers. ISBN 978-0-632-05614-9.
- ^ Paul, G.S. (). Predatory Dinosaurs of the World. New York: Simon and Schuster. pp. 248–250. ISBN 978-0-671-61946-6.
- ^ Clark J.M.; Maryanska T.; Barsbold R (). „Therizinosauroidea”. The Dinosauria (2d edition). University of California Press. pp. 151–164. ISBN 978-0-520-24209-8.
- ^ „Dromaeosauridae”. The Dinosauria (2d edition). University of California Press. . pp. 196–210. ISBN 978-0-520-24209-8.
- ^ Amiot, R.; Buffetaut, E.; Lecuyer, C.; Wang, X.; Boudad, L.; Ding, Z.; Fourel, F.; Hutt, S.; Martineau, F.; Medeiros, M.A.; Mo, J.; Simon, L.; Suteethorn, V.; Sweetman, S.; Tong, H.; Zhang, F.; Zhou, Z. (). „Oxygen isotope evidence for semi-aquatic habits among spinosaurid theropods”. Geology. 38 (2): 139–142. Bibcode:2010Geo....38..139A. doi:10.1130/G30402.1.
- ^ Taylor, M.P.; Wedel, M.J. (). „Why sauropods had long necks; and why giraffes have short necks”. PeerJ. 1: e36. doi:10.7717/peerj.36. PMC 3628838 . PMID 23638372.
- ^ a b Dal Sasso, C.; Signore, M. (). „Exceptional soft-tissue preservation in a theropod dinosaur from Italy”. Nature. 392 (6674): 383–387. Bibcode:1998Natur.392..383D. doi:10.1038/32884.
- ^ a b c Schweitzer, M.H.; Wittmeyer, J.L.; Horner, J.R. (). „Soft-tissue vessels and cellular preservation in Tyrannosaurus rex”. Science. 307 (5717): 1952–1955. Bibcode:2005Sci...307.1952S. doi:10.1126/science.1108397. PMID 15790853.
- ^ a b Alexander, R.M. (). „Dinosaur biomechanics”. Proceedings of the Royal Society B: Biological Sciences. 273 (1596): 1849–1855. doi:10.1098/rspb.2006.3532. PMC 1634776 . PMID 16822743.
- ^ Farlow, J.O.; Dodson, P.; Chinsamy, A. (). „Dinosaur Biology”. Annual Review of Ecology and Systematics. 26: 445–471. doi:10.1146/annurev.es.26.110195.002305.
- ^ Farlow JA (). „On the rareness of big, fierce animals: speculations about the body sizes, population densities, and geographic ranges of predatory mammals and large, carnivorous dinosaurs”. În Dodson, Peter; Gingerich, Philip. Functional Morphology and Evolution. American Journal of Science, Special Volume 293-A. pp. 167–199.
- ^ Peczkis, J. (). „Implications of body-mass estimates for dinosaurs”. Journal of Vertebrate Paleontology. 14 (4): 520–533. doi:10.1080/02724634.1995.10011575.
- ^ „Anatomy and evolution”. National Museum of Natural History. Accesat în .
- ^ Sander, P. Martin; Christian, Andreas; Clauss, Marcus; Fechner, Regina; Gee, Carole T.; Griebeler, Eva-Maria; Gunga, Hanns-Christian; Hummel, Jürgen; Mallison, Heinrich; et al. (). „Biology of the sauropod dinosaurs: the evolution of gigantism”. Biological Reviews. 86 (1): 117–155. doi:10.1111/j.1469-185X.2010.00137.x. PMC 3045712 . PMID 21251189.
- ^ Paul, Gregory S. (). Princeton Field Guide to Dinosaurs. Princeton University Press. ISBN 978-0-691-13720-9.
- ^ Colbert, Edwin Harris (). Men and dinosaurs: the search in field and laboratory. Harmondsworth [Eng.]: Penguin. ISBN 978-0-14-021288-4.
- ^ Mazzetta, G.V.; et al. (). „Giants and Bizarres: Body Size of Some Southern South American Cretaceous Dinosaurs” (PDF). Historical Biology. 16 (2–4): 1–13. doi:10.1080/08912960410001715132.
- ^ Janensch, W. (). „The Skeleton Reconstruction of Brachiosaurus brancai”: 97–103.
- ^ Lucas, H.; Hecket, H. (). „Reappraisal of Seismosaurus, a Late Jurassic Sauropod”. Proceeding, Annual Meeting of the Society of Paleontology. 36 (5): 422.
- ^ Sellers, W.I.; Margetts, L.; Coria, R.A.B.; Manning, P.L. (). Carrier, David, ed. „March of the Titans: The Locomotor Capabilities of Sauropod Dinosaurs”. PLOS One. 8 (10): e78733. Bibcode:2013PLoSO...878733S. doi:10.1371/journal.pone.0078733. PMC 3864407 . PMID 24348896.
- ^ Lovelace, David M. (). „Morphology of a specimen of Supersaurus (Dinosauria, Sauropoda) from the Morrison Formation of Wyoming, and a re-evaluation of diplodocid phylogeny”. Arquivos do Museu Nacional. 65 (4): 527–544.
- ^ „The fragile legacy of Amphicoelias fragillimus” (PDF). Arhivat din original (PDF) la . Accesat în .
- ^ „New information on the skull of the enigmatic theropod Spinosaurus, with remarks on its sizes and affinities” (PDF). Journal of Vertebrate Paleontology. 25 (4): 888–896. . doi:10.1671/0272-4634(2005)025[0888:NIOTSO]2.0.CO;2. Arhivat din original (PDF) la . Accesat în .
- ^ a b Therrien, F.; Henderson, D.M. (). „My theropod is bigger than yours ... or not: estimating body size from skull length in theropods”. Journal of Vertebrate Paleontology. 27 (1): 108–115. doi:10.1671/0272-4634(2007)27[108:MTIBTY]2.0.CO;2.
- ^ Norell, M., Gaffney, E.S., and Dingus, L. (2000). Discovering dinosaurs: Evolution, extinction, and the lessons of prehistory. University of California Press.
- ^ „Bee Hummingbird (Mellisuga helenae) : Birds.com: Online Birds Guide with Facts, Articles, Videos, and Photos”. Arhivat din original la . Accesat în .
- ^ a b „A bizarre Jurassic maniraptoran from China with elongate ribbon-like feathers”. Nature. 455 (7216): 1105–1108. . Bibcode:2008Natur.455.1105Z. doi:10.1038/nature07447. PMID 18948955.
- ^ a b „A new feathered maniraptoran dinosaur fossil that fills a morphological gap in avian origin”. Chinese Science Bulletin. 54 (3): 430–435. . doi:10.1007/s11434-009-0009-6.
- ^ Butler, R.J.; Zhao, Q. (). „The small-bodied ornithischian dinosaurs Micropachycephalosaurus hongtuyanensis and Wannanosaurus yansiensis from the Late Cretaceous of China”. Cretaceous Research. 30 (1): 63–77. doi:10.1016/j.cretres.2008.03.002.
- ^ Yans J; Dejax J; Pons D; Dupuis C; Taquet P (). „Implications paléontologiques et géodynamiques de la datation palynologique des sédiments à faciès wealdien de Bernissart (bassin de Mons, Belgique)”. Comptes Rendus Palevol (în French). 4 (1–2): 135–150. doi:10.1016/j.crpv.2004.12.003.
- ^ Deposition of Iguanodon skeletons occurred in at least 3 different events. Bernissart Iguanodons and their significance (long) date : 4 Jul. 2001.
- ^ Wright, Joanna L. (). „Steps in understanding sauropod biology”. În Curry Rogers; Kristina A.; Wilson, Jeffrey A. The Sauropods: Evolution and Paleobiology. Berkeley: University of California Press. pp. 252–284. ISBN 978-0-520-24623-2.
- ^ Varricchio, D.J.; Sereno, Paul C.; Xijin, Zhao; Lin, Tan; Wilson, Jeffery A.; Lyon, Gabrielle H. (). „Mud-trapped herd captures evidence of distinctive dinosaur sociality” (PDF). Acta Palaeontologica Polonica. 53 (4): 567–578. doi:10.4202/app.2008.0402. Accesat în .
- ^ Lessem, Don; Glut, Donald F. (). „Allosaurus”. The Dinosaur Society's Dinosaur Encyclopedia. Random House. pp. 19–20. ISBN 978-0-679-41770-5.
- ^ a b Maxwell, W.D.; Ostrom, John (). „Taphonomy and paleobiological implications of Tenontosaurus–Deinonychus associations”. Journal of Vertebrate Paleontology. 15 (4): 707–712. doi:10.1080/02724634.1995.10011256.
- ^ Roach, Brian T.; Brinkman, Daniel L. (). „A reevaluation of cooperative pack hunting and gregariousness in Deinonychus antirrhopus and other nonavian theropod dinosaurs”. Bulletin of the Peabody Museum of Natural History. 48 (1): 103–138. doi:10.3374/0079-032X(2007)48[103:AROCPH]2.0.CO;2.
- ^ Tanke, Darren H. (). „Head-biting behavior in theropod dinosaurs: paleopathological evidence” (PDF). Gaia (15): 167–184. Arhivat din original (PDF) la .
- ^ „The Fighting Dinosaurs”. American Museum of Natural History. Arhivat din original la . Accesat în .
- ^ a b Carpenter, K. (). „Evidence of predatory behavior by theropod dinosaurs” (PDF). Gaia. 15: 135–144.
- ^ Rogers, Raymond R.; Krause, DW; Curry Rogers, K (). „Cannibalism in the Madagascan dinosaur Majungatholus atopus”. Nature. 422 (6931): 515–518. Bibcode:2003Natur.422..515R. doi:10.1038/nature01532. PMID 12673249.
- ^ „Dinosaur babies took a long time to break out of their shells”..
- ^ Schmitz, L.; Motani, R. (). „Nocturnality in Dinosaurs Inferred from Scleral Ring and Orbit Morphology”. Science. 332 (6030): 705–708. Bibcode:2011Sci...332..705S. doi:10.1126/science.1200043. PMID 21493820.
- ^ a b Senter, P. (). „Voices of the past: a review of Paleozoic and Mesozoic animal sounds”. Historical Biology. 20 (4): 255–287. doi:10.1080/08912960903033327.
- ^ Li, Q.; et al. (). „Plumage Color Patterns of an Extinct Dinosaur” (PDF). Science. 327 (5971): 1369–1372. Bibcode:2010Sci...327.1369L. doi:10.1126/science.1186290. PMID 20133521.
- ^ Clarke, J.A.; et al. (). „Fossil evidence of the avian vocal organ from the Mesozoic”. Nature. 538 (7626): 502–505. Bibcode:2016Natur.538..502C. doi:10.1038/nature19852. PMID 27732575.
- ^ Weishampel, D.B. (). „Acoustic Analysis of Vocalization of Lambeosaurine Dinosaurs (Reptilia: Ornithischia)” (PDF). Paleobiology. 7 (2): 252–261. Arhivat din original (PDF) la .
- ^ Miyashita, T.; Arbour V.M.; Witmer L.M.; Currie, P.J. (). „The internal cranial morphology of an armoured dinosaur Euoplocephalus corroborated by X-ray computed tomographic reconstruction” (PDF). Journal of Anatomy. 219 (6): 661–675. doi:10.1111/j.1469-7580.2011.01427.x. PMC 3237876 . PMID 21954840. Arhivat din original (PDF) la .
- ^ Currie, Philip J; Kevin Padian, ed. (). Encyclopedia of Dinosaurs. Academic Press. p. 206.
- ^ Hansell M (2000). Bird Nests and Construction Behaviour. University of Cambridge Press ISBN: 0-521-46038-7
- ^ Varricchio, David J.; Horner, John J.; Jackson, Frankie D. (). „Embryos and eggs for the Cretaceous theropod dinosaur Troodon formosus”. Journal of Vertebrate Paleontology. 22 (3): 564–576. doi:10.1671/0272-4634(2002)022[0564:EAEFTC]2.0.CO;2.
- ^ Lee, Andrew H.; Werning, S (). „Sexual maturity in growing dinosaurs does not fit reptilian growth models”. Proceedings of the National Academy of Sciences. 105 (2): 582–587. Bibcode:2008PNAS..105..582L. doi:10.1073/pnas.0708903105. PMC 2206579 . PMID 18195356.
- ^ a b Horner, J.R.; Makela, Robert (1979). "Nest of juveniles provides evidence of family structure among dinosaurs". Nature 282 (5736): 296–298. Bibcode:1979Natur.282..296H. doi:10.1038/282296a0
- ^ Reisz, RR; Scott, D; Sues, H-D; Evans, DC; Raath, MA (). „Embryos of an Early Jurassic prosauropod dinosaur and their evolutionary significance” (PDF). Science (Submitted manuscript). 309 (5735): 761–764. Bibcode:2005Sci...309..761R. doi:10.1126/science.1114942. PMID 16051793.
- ^ „Dinosaur footprints from the Duntulm Formation (Bathonian, Jurassic) of the Isle of Skye” (PDF). Scottish Journal of Geology. 40 (1): 13–21. doi:10.1144/sjg40010013. Accesat în .
- ^ Huxley, Thomas H. (). „On the animals which are most nearly intermediate between birds and reptiles”. The Annals and Magazine of Natural History. 4 (2): 66–75.
- ^ a b Heilmann, Gerhard (). The Origin of Birds. London: Witherby. pp. 208pp. ISBN 978-0-486-22784-9.
- ^ Osborn, Henry Fairfield (). „Three new Theropoda, Protoceratops zone, central Mongolia” (PDF). American Museum Novitates. 144: 1–12.
- ^ Ostrom, John H. (). „The ancestry of birds”. Letters to Nature. 242 (5393): 136. doi:10.1038/242136a0.
- ^ Gauthier, Jacques (). „Saurischian monophyly and the origin of birds”. În Padian, Kevin. The Origin of Birds and the Evolution of Flight. Memoirs of the California Academy of Sciences 8. pp. 1–55.
- ^ Holtz Jr., T.R. (). „Classification and evolution of the dinosaur groups”. În Paul, G.S. The Scientific American Book of Dinosaurs. St. Martin's Press. pp. 140–168. ISBN 978-0-312-26226-6.
- ^ Mayr, G.; Pohl, B.; Peters, D.S. (). „A Well-Preserved Archaeopteryx Specimen with Theropod Features”. Science. 310 (5753): 1483–1486. Bibcode:2005Sci...310.1483M. doi:10.1126/science.1120331. PMID 16322455.
- ^ Martin, Larry D. (). „A basal archosaurian origin for birds”. Acta Zoologica Sinica. 50 (6): 977–990.
- ^ a b Feduccia, A. (). „Birds are dinosaurs: simple answer to a complex problem”. The Auk. 119 (4): 1187–1201. doi:10.1642/0004-8038(2002)119[1187:BADSAT]2.0.CO;2.
- ^ Switek, B. (). „Rise of the fuzzy dinosaurs”. Nature. doi:10.1038/nature.2012.10933.
- ^ Godefroit, P., Sinitsa, S., Dhouailly, D., Bolotsky, Y., and Sizov, A. "Feather-like structures and scales in a Jurassic neornithischian dinosaur from Siberia." Program and Abstracts of the 73rd Meeting of the Society of Vertebrate Paleontology Arhivat în , la Wayback Machine., October 2013.
- ^ Xu X.; Norell, M.A.; Kuang X.; Wang X.; Zhao Q.; Jia C. (). „Basal tyrannosauroids from China and evidence for protofeathers in tyrannosauroids”. Nature. 431 (7009): 680–684. Bibcode:2004Natur.431..680X. doi:10.1038/nature02855. PMID 15470426.
- ^ Göhlich, U.B.; Chiappe, LM (). „A new carnivorous dinosaur from the Late Jurassic Solnhofen archipelago” (PDF). Nature. 440 (7082): 329–332. Bibcode:2006Natur.440..329G. doi:10.1038/nature04579. PMID 16541071.
- ^ Kellner, A.W.A.; Wang, X.; Tischlinger, H.; Campos, D.; Hone, D.W.E.; Meng, X. (). „The soft tissue of Jeholopterus (Pterosauria, Anurognathidae, Batrachognathinae) and the structure of the pterosaur wing membrane”. Proceedings of the Royal Society B. 277 (1679): 321–329. doi:10.1098/rspb.2009.0846. PMC 2842671 . PMID 19656798.
- ^ a b Wellnhofer, P (). „Ein neuer Exemplar von Archaeopteryx”. Archaeopteryx. 6: 1–30.
- ^ a b Schweitzer, M.H.; Watt, J.A.; Avci, R.; Knapp, L.; Chiappe, L.; Norell, M.; Marshall, M. (). „Beta-keratin specific immunological reactivity in feather-like structures of the Cretaceous Alvarezsaurid, Shuvuuia deserti”. Journal of Experimental Zoology. 285 (2): 146–157. doi:10.1002/(SICI)1097-010X(19990815)285:2<146::AID-JEZ7>3.0.CO;2-A. PMID 10440726.
- ^ Lingham-Soliar, T. (). „The dinosaurian origin of feathers: perspectives from dolphin (Cetacea) collagen fibers”. Naturwissenschaften. 90 (12): 563–567. Bibcode:2003NW.....90..563L. doi:10.1007/s00114-003-0483-7. PMID 14676953.
- ^ a b Feduccia, A.; Lingham-Soliar, T; Hinchliffe, JR (). „Do feathered dinosaurs exist? Testing the hypothesis on neontological and paleontological evidence”. Journal of Morphology. 266 (2): 125–166. doi:10.1002/jmor.10382. PMID 16217748.
- ^ Lingham-Soliar, T.; Feduccia, A; Wang, X (). „A new Chinese specimen indicates that 'protofeathers' in the Early Cretaceous theropod dinosaur Sinosauropteryx are degraded collagen fibres”. Proceedings of the Royal Society B. 274 (1620): 1823–1829. doi:10.1098/rspb.2007.0352. PMC 2270928 . PMID 17521978.
- ^ Prum, Richard O. (aprilie 2003). „Are Current Critiques Of The Theropod Origin Of Birds Science? Rebuttal To Feduccia 2002”. The Auk. 120 (2): 550–561. doi:10.1642/0004-8038(2003)120[0550:ACCOTT]2.0.CO;2.
- ^ „First Dinosaur Tail Found Preserved in Amber”. . Accesat în .
- ^ Xing, L; McKellar, RC; Xu, X; Li, G; Bai, M; Persons; Miyashita, T; Benton, MJ; Zhang, J; Wolfe, AP; Yi, Q; Tseng, K; Ran, H; Currie, PJ (). „A Feathered Dinosaur Tail with Primitive Plumage Trapped in Mid-Cretaceous Amber”. Current Biology. 26 (24): 3352–3360. Bibcode:1996CBio....6.1213A. doi:10.1016/j.cub.2016.10.008. PMID 27939315.
- ^ Archaeopteryx: a missing link. Berkeley: University of California. Museum of Paleontology.
- ^ Dong Zhiming (). Dinosaurian Faunas of China. China Ocean Press, Beijing. ISBN 978-3-540-52084-9. OCLC 26522845.
- ^ Benton, M.J. (2000). "A brief history of dinosaur paleontology". pp. 10–44, In Paul, G.S. (ed.). The Scientific American book of dinosaurs. St. Martin's Press, New York.
- ^ a b c Sarjeant WAS (). „The earliert discoveries”. The Complete Dinosaur. Bloomington: Indiana University Press. pp. 3–11. ISBN 978-0-253-33349-0.
- ^ Lhuyd, E. (1699). Lithophylacii Britannici Ichnographia, sive lapidium aliorumque fossilium Britannicorum singulari figura insignium. Gleditsch and Weidmann:London.
- ^ Delair, J.B.; Sarjeant, W.A.S. (). „The earliest discoveries of dinosaurs: the records re-examined”. Proceedings of the Geologists' Association. 113 (3): 185–197. doi:10.1016/S0016-7878(02)80022-0.
- ^ a b Buckland W (). „Notice on the Megalosaurus or great Fossil Lizard of Stonesfield”. Transactions of the Geological Society of London. 1 (2): 390–396. doi:10.1144/transgslb.1.2.390.
- ^ a b Mantell, Gideon A. (). „Notice on the Iguanodon, a newly discovered fossil reptile, from the sandstone of Tilgate forest, in Sussex”. Philosophical Transactions of the Royal Society. 115: 179–186. doi:10.1098/rstl.1825.0010.
- ^ a b Sues, Hans-Dieter (). „European Dinosaur Hunters”. În Farlow JO; Brett-Surman MK. The Complete Dinosaur. Bloomington: Indiana University Press. p. 14. ISBN 978-0-253-33349-0.
- ^ a b Rupke, N. (1994). Richard Owen: A Victorian Naturalist. New Haven: Yale University Press.
- ^ Prieto-Marques, A.; Weishampel, D.B.; Horner, J.R. (). „The dinosaur Hadrosaurus foulkii, from the Campanian of the East Coast of North America, with a reevaluation of the genus”. Acta Palaeontologica Polonica. 51 (1): 77–98.
- ^ Holmes T (aprilie 1996). Fossil Feud: The Bone Wars of Cope and Marsh, Pioneers in Dinosaur Science. Silver Burdett Press. ISBN 978-0-382-39147-7. OCLC 34472600.
- ^ Salgado, L.; Gasparini, Z. (). „Reappraisal of an ankylosaurian dinosaur from the Upper Cretaceous of James Ross Island (Antarctica)”. Geodiversitas. 28 (1): 119–135.
- ^ Evershed, Nick (). „Blood, tissue extracted from duck-billed dinosaur bone”. Cosmosmagazine.com. Arhivat din original la . Accesat în .
- ^ „Molecular analyses of dinosaur osteocytes support the presence of endogenous molecules”. Bone. 52 (1): 414–423. ianuarie 2013. doi:10.1016/j.bone.2012.10.010. PMID 23085295.
- ^ Wang, H.; Yan, Z.; Jin, D. (). „Reanalysis of published DNA sequence amplified from Cretaceous dinosaur egg fossil”. Molecular Biology and Evolution. 14 (5): 589–591. doi:10.1093/oxfordjournals.molbev.a025796. PMID 9159936. Arhivat din original la . Accesat în .
- ^ „Recreating a Functional Ancestral Archosaur Visual Pigment”. Molecular Biology and Evolution. 19 (9): 1483–1489. . doi:10.1093/oxfordjournals.molbev.a004211. PMID 12200476. Accesat în .
- ^ „Heme compounds in dinosaur trabecular bone”. Proc Natl Acad Sci U S A. 94 (12): 6291–6296. . Bibcode:1997PNAS...94.6291S. doi:10.1073/pnas.94.12.6291. PMC 21042 . PMID 9177210.
- ^ „Identification of proteinaceous material in the bone of the dinosaur Iguanodon”. Connect Tissue Res. 44 (Suppl 1): 41–46. . doi:10.1080/03008200390152070. PMID 12952172.
- ^
Peterson, JE; Lenczewski, ME; Reed, PS (octombrie 2010). Stepanova, Anna, ed. „Influence of Microbial Biofilms on the Preservation of Primary Soft Tissue in Fossil and Extant Archosaurs”. PLOS One. 5 (10): 13A. Bibcode:2010PLoSO...513334P. doi:10.1371/journal.pone.0013334. PMC 2953520 . PMID 20967227.
- ^ Bertazzo, Sergio; Maidment, Susannah C.R.; Kallepitis, Charalambos; Fearn, Sarah; Stevens, Molly M.; Xie, Hai-nan (). „Fibres and cellular structures preserved in 75-million-year-old dinosaur specimens”. Nature Communications. 6: 7352. Bibcode:2015NatCo...6E7352B. doi:10.1038/ncomms8352. PMC 4468865 . PMID 26056764.
- ^ Mortillaro, Nicole (). „Scientists discover 75-million-year-old dinosaur blood and tissue”. Accesat în .
Legături externe
modificareGeneral
Images
- The Science and Art of Gregory S. Paul Influential paleontologist's anatomy art and paintings
- Skeletal Drawing Professional restorations of numerous dinosaurs, and discussions of dinosaur anatomy.
- "Dinosaur Discovery" Arhivat în , la Wayback Machine., a collection of images from early works on dinosaurs at the Linda Hall Library, in support of the exhibition, Paper Dinosaurs, 1824–1969 Arhivat în , la Wayback Machine..
Video
- BBC Nature: Dinosaur reconstructions and expert interpretations, including Walking with Dinosaurs
- BBC Explainer – Dinosaurs – a complete history in 4 minutes, animation
- Origin, evolution and extinction of the dinosaurs Stephen Brusatte video lecture 15 aprilie 2014